INTRODUCTION
Liver regeneration has been extensively studied [1-3]. In vivo studies have shown that partial hepatectomy or chemical injury activates extracellular and intracellular signaling pathways, leading to liver regeneration. Hepatocyte loss during chronic liver diseases triggers compensatory proliferation of the surviving hepatocytes [4-6]. Apart from liver regeneration in physiological conditions, genotoxic risk factors might lead them to convert to neoplasia. Hepatitis virus, alcohol abuse, non-alcoholic fatty liver disease (NAFLD), and aflatoxin-B1 exposure are also the main etiological factors to induce the development of precancerous lesions in the liver. Liver cancer is one of the top 10 lethal cancers worldwide. Its estimated death rate in 2021 is 6% in males and 4% in females [7]. Liver cancer consists of hepatocellular carcinoma (HCC), cholangiocarcinoma (CCA), hepatoblastoma (HB), and several other rare tumors (angiosarcoma, intraductal papillary neoplasm of the bile duct, and mucinous cystic neoplasm). HCC is the most common primary liver cancer frequently developed with chronic liver disease, such as cirrhosis caused by hepatitis virus infection [8].
Among various signaling pathways associated with liver biology [9-12], Wnt signaling is involved in all stages of liver disease progression, from liver injury to inflammation, fibrosis, cirrhosis, and tumorigenesis. Several Wnt ligands are secreted by various hepatic cells, including hepatocytes, stellate cells, Kupffer cells, biliary epithelial cells, and sinusoidal endothelial cells [13-16]. Based on the oncogenic roles of Wnt signaling in cancer, several components and regulators of Wnt signaling have been proposed as the druggable targets to improve the current therapeutic efficacy in the liver cancer treatment [17].
Herein, we review the roles of Wnt signaling in liver regeneration and liver tumorigenesis and the therapeutic targets of Wnt signaling in liver cancer treatment.
Wnt SIGNALING
Wnt signaling is evolutionarily conserved and orchestrates various cellular processes, including cell proliferation, differentiation, migration, polarity, stemness, and lineage plasticity [18,19]. Consequently, Wnt signaling plays a pivotal role in organogenesis, tissue homeostasis, tissue regeneration, and tumorigenesis [20-25]. The Wnt signaling is triggered by the binding of the Wnt ligands to the frizzed (FZD) receptors. The mammals have 19 Wnt ligands and 10 FZD receptors [26], resulting in the complexity and specificity in Wnt signaling activation. Based on the involvement of β-catenin, a key component of Wnt signaling, Wnt signaling is generally classified into canonical (β-catenin-mediated) and non-canonical (β-catenin-independent) Wnt signaling (Fig. 1). In the canonical Wnt/β-catenin pathway, the protein destruction complex (casein kinase 1 [CK1], glycogen synthase kinase 3 [GSK3], adenomatous polyposis coli [APC], and axis inhibition proteins [AXINs]) targets the β-catenin protein for degradation via CKI1 and GSK3-mediated sequential phosphorylation at the N-terminus (Ser-45, Thr-41, Ser-37, and Ser-33) of β-catenin followed by β-TrCP, an E3 ligase, recruitment. Conversely, binding of the canonical Wnt ligands to the FZD receptors and LRP5/6 co-receptors activates dishevelled (DVL), which inhibits the protein destruction complex. As a result, β-catenin protein is stabilized and translocated into the nucleus to transactivate the canonical Wnt target genes by replacing the co-repressors associated with the T-cell factor/lymphoid enhancer-binding factor (TCF/LEF) with the co-activators. Non-canonical Wnt signaling pathways include the planar cell polarity pathway (involved in c-Jun N-terminal kinase [JNK] activation, small GTPase activation, and cytoskeletal rearrangement), and the Wnt/Ca2+ pathway (activating phospholipase C [PLC] and protein kinase C [PKC]) [18,19].
Wnt SIGNALING IN LIVER REGENERATION
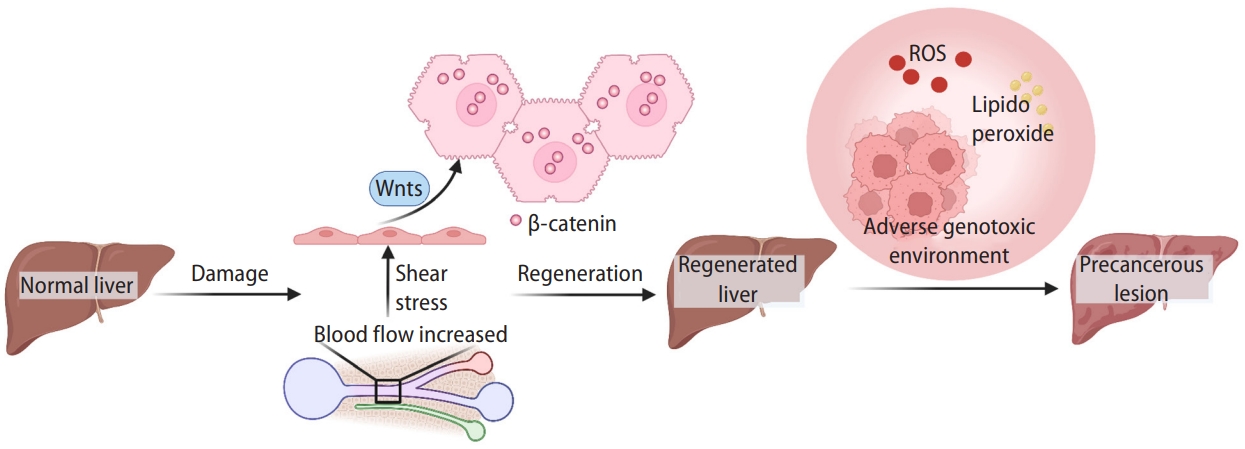
Upon partial hepatectomy or acute liver injury, the number of hepatocytes is drastically reduced. Various signaling pathways (epidermal growth factor [EGF], hepatocyte growth factor [HGF], Wnt/β-catenin, and Notch) stimulate the hepatocytes in the G0 phase to proliferate, compensating tissue loss and restoring the physiological functions of the liver [27-29]. During liver regeneration, endothelial cells under shear stress produce Wnts to activate Wnt/β-catenin signaling in hepatocytes. Additionally, the organ precisely senses the size of the regenerating liver and adjusts its size to 100% [2].
Several animal models (rat, mouse, and zebrafish) were utilized for liver regeneration study [30-33]. The partial hepatectomy is the classic strategy to create the murine liver regeneration model [30]. Carbon tetrachloride (CCl4)is a frequently used chemical to induce liver injury in rats and mice.34 Meanwhile, several dietary-induced liver injury models are also commonly used [35]. Biliary injury and regeneration can be induced by the 1,4-dihydro-2,4,6-trimethyl-pyridine-3,5-dicarboxylate (DDC) diet [35]. Besides murine models, zebrafish emerged as a potent model for drug screening of liver generation [32,33]. Partial hepatectomy, drug-induced liver injury, and nitroreductase-mediated hepatocyte ablation were employed to establish the zebrafish liver injury model [32,33,36,37].
Transient activation of the canonical Wnt signaling is indispensable for liver regeneration (Fig. 2) [13,15,27]. In rat models, overexpressed Wnt1 and nuclear β-catenin are predominantly accumulated in remaining parenchymal cells after 70% partial hepatectomy. The level of β-catenin increased within 5 minutes after hepatectomy, accompanied by its nuclear translocation and subsequent target gene expression for hepatocyte proliferation [27]. Significantly, genetic ablation of β-catenin/Ctnnb1 impairs liver regeneration of mice from partial hepatectomy [38]. The liver-specific Ctnnb1 knock-out (KO) delayed DNA synthesis and hepatocyte proliferation in mice after partial hepatectomy. Conversely, activation of Wnt/β-catenin signaling accelerates liver regeneration in the zebrafish model [39]. It was also shown that liver damage upregulated leucine-containing repeat G-protein-coupled receptor 5 (LGR5) and AXIN2 in the hepatocytes [40]. LGR5 is a marker of actively dividing stem and progenitor cells in Wnt-driven self-renewing tissues [41]. LGR5 interacts with FZD and lipoprotein receptor-related protein 6 (LRP6) to enhance phosphorylation of LRP6, which in turn enhances the Wnt/β-catenin signaling [42]. While Lgr5 is not expressed in healthy adult livers, after liver damage, Lgr5+ cells appear near the bile ducts, consistent with strong activation of the Wnt signaling [41]. AXIN2 is another Wnt downstream target gene transactivated by β-catenin [43]. Like AXIN1, AXIN2 combined with other destruction complex components degrades β-catenin, serving as a negative feedback regulator of the Wnt signaling [44].
Other than core components of Wnt signaling, additional regulators of Wnt signaling were implicated in liver regeneration. Recently, our group identified the transmembrane protein 9 (TMEM9) gene as an amplifier of Wnt/β-catenin signaling. TMEM9 is a type I transmembrane protein primarily localized in lysosomes and multivesicular bodies (MVBs). While the ablation of TMEM9 inhibits the activity of the Wnt/β-catenin signaling, β-catenin transactivates TMEM9, leading to hyperactivation of Wnt/β-catenin signaling [45]. Interestingly, TMEM9 is highly expressed in hepatocytes around the central vein (CV) of regenerating liver [46]. TMEM9 hyperactivates Wnt/β-catenin signaling to promote liver regeneration through lysosomal degradation of APC protein [46]. Tmem9 KO impairs CCl4-induced liver regeneration with downregulation of Wnt/β-catenin signaling [46].
In addition to the role of Wnt/β-catenin signaling in regeneration, sustained activation of the Wnt signaling is associated with the progression of chronic liver diseases and liver tumorigenesis (Fig. 2). Additionally, reactive oxygen species (ROS) and lipid peroxide are the risk factors for the development of the precancerous lesion in the liver [47,48]. However, the crosstalk between Wnt signaling and ROS has not been fully revealed in the liver. It was reported that β-catenin can be further stabilized by ROS [49]. Meanwhile, lipid peroxidation products mainly generated by ROS activate the canonical Wnt pathway through oxidative stress [50]. Therefore, it is likely the potential crosstalk between Wnt signaling and ROS might contribute to liver cancer development.
Accumulating evidence suggests that many chronic liver diseases contribute to liver cancer development, described below.
Wnt SIGNALING IN PRECANCEROUS LIVER LESION
Hepatitis virus
Globally distributed hepatitis B virus (HBV) and hepatitis C virus (HCV) are the crucial triggers of HCC initiation. Both HBV and HCV can induce chronic infections and are essential pathogenic factors in cirrhosis and liver cancer (Fig. 3) [51,52]. The epidemiological data show that more than 70% of patients with liver cancer have HBV infection, 10–20% have HCV infection, and a significant proportion of patients have both HBV and HCV infection [53-55].
After infection, the DNA of HBV is integrated into the host genome, inducing genomic instability and transactivation of cancer-related genes, which culminates in the formation of early cancer cell clones. Mechanistically, HBV contributes to HCC development through direct and indirect means [56]. Direct mechanisms include virus mutations, HBV DNA integration, growth regulatory genes activation by HBV-encoded proteins [57]. Indirect mechanisms include the activation of cellular oncogenes associated with HBV DNA integration, genetic instability induced by viral integration or the regulatory protein HBx, and the development of liver disease mediated by immune enhancement due to viral proteins [58].
Both hepatitis B virus surface antigen (HBsAg) and HBx modulate the expressions of genes involved in Wnt signaling activation. HBsAg activates the transcription factor LEF1 of the Wnt signaling [59]. The X protein encoded by the hepatitis B virus has a vital role in stimulating viral gene expression and replication, critical for maintaining chronic carrier status. HBx, a 17 kDa multifunctional protein, upregulates the expression of Wnt ligands (WNT1 and WNT3), the receptor (FZD2 and FZD7), a component of the destruction complex (GSK3β), E-cadherin, and Wnt1-inducible-signaling pathway protein 1 (WISP1), a suppressor of Wnt antagonists (secreted frizzled-related protein 1 [SFRP1] and SFRP5). On the other hand, the Wnt signaling key components (β-catenin and AXIN1) are highly mutated in HBV-associated HCC. Loss-of-function (LOF) mutations of the AXIN1 are observed in HBV-HCC patients. In HBV and/or HCV-associated HCC patients, the most frequent mutation in the CTNNB1 gene is enriched in the exon 3 encoding the N-terminal phosphorylation sites [60-62]. These aberrantly controlled genes in Wnt signaling subsequently promote and lead to the development of HCC [63-65].
The oncogenic mechanism of HCV in liver cancer is mainly mediated by Wnt/β-catenin signaling hyperactivation via the core protein and two nonstructural proteins, NS3 and NS5A [66]. The core protein (HCV core antigen) is a significant component of HCV. It regulates hepatocyte transcription and promotes Wnt/β-catenin signaling by upregulating Wnt ligands (WNT1 and WNT3A), FZD receptors, and LRP5/6 [67,68]. Additionally, at the early stage of HCV infection, the secreted Wnt antagonists, SFRP2 and Dickkopf 1 (DKK1), are downregulated by their promoter hypermethylation [69,70]. HCV core protein also promotes hypermethylation of the CDH1 gene promoter region [71], destabilizing the cadherin-catenin-actin complex for β-catenin release and activation [72]. NS5A stabilizes β-catenin via activating phosphoinositide 3-kinase (PI3K)/AKT, leading to GSK3β inactivation followed by inhibiting the protein-destruction complex-mediated β-catenin degradation for Wnt target gene activation. At the early stage of viral infection, HCV-activated Wnt/β-catenin signaling also promotes liver fibrosis by enhancing the activation and survival of hepatic stellate cells [17,73,74].
Alcohol abuse
Alcohol is a well-known risk factor for liver cancer. Alcoholic liver disease (ALD) is a chronic liver disease caused by long-term alcohol consumption (Fig. 3). ALD is characterized by the fatty liver at the beginning, then progressed to alcoholic hepatitis, liver fibrosis, and cirrhosis, which is pathologically associated with the precancerous lesions of HCC. In vivo, ethanol (EtOH) is metabolized into the reactive metabolite acetaldehyde, promoting liver tumorigenesis. Mice administered with the chemical carcinogen, diethylnitrosamine (DEN), for 7 weeks and the subsequent EtOH feeding for 16 weeks exhibited the increased total number of cancer foci and liver tumors [75]. Also, these tumors showed a 3- to 4-fold increase in the expression of proliferation markers and an increased expression of β-catenin, compared to non-tumor hepatocytes [75]. In a rat model of chronic liver disease, EtOH-treated liver was accompanied by the increased proliferation of hepatocytes, depletion of retinol and retinoic acid storage, augmented expression of phospho-GSK3β at the cell membrane, significant upregulation of soluble Wnt ligands (Wnt2 and Wnt7a), accumulation of nuclear β-catenin, and upregulation of β-catenin target genes (cyclin D1/CCND1, c-Myc/MYC, WISP1, and matrix metallopeptidase [MMP7]). These data suggest that long-term EtOH consumption activates the Wnt/β-catenin signaling and increases hepatocyte proliferation, promoting liver tumorigenesis [75]. Additionally, ROS accumulation, nuclear factor kappa-light-chain-enhancer of activated B cells (NF-ĸB)-dependent vascular endothelial growth factor (VEGF) and monocyte chemoattractant protein (MCP)-1 upregulation, and activation of extracellular signal-regulated kinase (ERK)-mitogen-activated protein kinase (MAPK) signaling also contribute to EtOH-induced liver tumorigenesis [76-80].
NAFLD
The increasing prevalence of NAFLD was caused by an over-nourished lifestyle [81,82]. NAFLD is characterized by fat accumulation in the liver, evolving to end-stage liver diseases such as cirrhosis and HCC (Fig. 3) [83]. The main risk factors of NAFLD include central obesity, overnutrition, insulin resistance, and metabolic syndrome [84]. In severe NAFLD, many tissue repair-related genes (TMEM204, FGFR2, matrix molecules, and matrix remodeling factors) were hypomethylated at their promoters and overexpressed. Conversely, genes in specific metabolic pathways (lipid metabolism, cytochrome P450 family, multidrug resistance, and fatty acid anabolic pathways) were hypermethylated and silenced [85]. Hyperinsulinemia is one of the risk factors of NAFLD [86]. SOX17 plays a vital role in regulating insulin secretion. Sox17 KO mice display high susceptibility to high-fat diet-induced hyperglycemia and diabetes [87]. SOX17 directly interacts with the TCF/LEF transcription factor to repress the transcription of Wnt signaling target genes. The methylation of the SOX17 promoter is a frequent event in human cancers. Epigenetic silencing of SOX17 contributes to the aberrant activation of Wnt/β-catenin signaling [88], accelerating progression from NAFLD to HCC. Besides, β-catenin inhibits the expression of CCAAT enhancer-binding protein α (CEBPA) and peroxisome proliferator-activated receptor γ (PPARG), which in turn inhibits the preadipocyte differentiation [89]. As the co-receptor of the Wnt/β-catenin signaling, LRP6 induces lipid accumulation in the liver via insulin-like growth factor 1 (IGF1)/AKT/mammalian target of rapamycin (mTOR)/sterol regulatory element binding transcription factor (SREBF) 1/2 signaling. Intriguingly, inhibiting the non-canonical Wnt signaling reduces lipid accumulation and inflammation [90]. Therefore, while reducing the effects of NAFLD risk factors, inhibition of the Wnt signaling is also essential for attenuating the development of NAFLD and preventing the initiation of HCC.
Aflatoxin-B1 exposure
Among the aflatoxins, aflatoxin type B1 (AFB1) primarily targets the liver as a highly potent hepatotoxin and hepatocarcinogen (Fig. 3). AFB1 impairs DNA repair processes, resulting in severe DNA mutagenesis, and also inhibits DNA and RNA metabolism. This pathological event ultimately leads to excessive liver lipid accumulation, liver enlargement, bile duct epithelial hyperplasia, and liver cancer. The potency of aflatoxin to cause liver cancer is significantly enhanced in the presence of HBV infection. Under chronic HBV infection, cytochrome P450s could metabolize inactive AFB1 to mutagenic AFB1-8,9-epoxide. Also, the infection leads to hepatocyte necrosis and regeneration, producing oxygen and nitrogen reactive species and increasing the incidence of AFB1-induced mutagenesis [91]. Clinical studies have shown that CTNNB1 mutations are present in approximately one-quarter of HCC in areas with low aflatoxin B1 exposure. Interestingly, these CTNNB1 mutations were similar to those previously reported in the human HCC [92].
Wnt SIGNALING IN LIVER CANCER
HCC
HCC is a common and fatal malignancy worldwide [93]. Regardless of the risk factors mentioned above, aberrant hyperactivation of Wnt/β-catenin signaling is observed in 95% of HCCs [94]. The most common genetic mutations of the Wnt signaling in HCC are the gain-of-function mutations in the CTNNB1 gene encoding β-catenin [61,95], which is somewhat distinct from colorectal cancer where Wnt/β-catenin signaling hyperactivation is mainly driven by the APC gene inactivation [96]. Missense mutations of CTNNB1 exon 3 were observed in 18.1% of HCC cases. Missense mutations at codons 32, 33, 38, or 45 of the CTNNB1 gene lead to the unphosphorylation of the N-terminus of β-catenin for its stabilization, nuclear translocation, and target gene transactivation [60]. Secondly, the LOF mutations in the AXIN1 gene were observed in 5–19% of HCC cases [97]. CTNNB1 and AXIN1 mutations occur in patients with advanced HCC (Fig. 3) [98-100]. Importantly, hyperactivation of the Wnt signaling is considered a hallmark of advanced HCC [101]. It should also be noted that mutations in the CTNNB1 and AXIN1 genes lead to different HCC subtypes accompanied by distinct clinical and pathological features. CTNNB1 mutations are associated with less aggressive HCC, including chromosomally stable and highly differentiated tumors [102], with a better prognosis [95]. In contrast, AXIN1 mutations occur more frequently in more aggressive HCC tumors characterized by hypodifferentiated tumor cells and chromatin instability [102]. Consistently, the HCC tumors with CTNNB1 mutations or AXIN1 mutations showed different target gene expression [61,95,103].
CCA
CCA is ranked as the second most common hepatobiliary cancer after HCC. CCA originates mainly from differentiated bile duct epithelial cells [104]. CCA is often diagnosed at an advanced stage with a poor prognosis. Current chemotherapy has not improved the survival rate of unresectable CCA patients. Clinical and preclinical studies have shown that activation of the Wnt/β-catenin signaling occurs throughout the initiation and progression of CCA. Wnt ligands (WNT2, WNT7b, and WNT10A) and TCF4 are upregulated in CCA, accompanied by nuclear translocation of β-catenin [105,106]. The progression of epithelial-mesenchymal transition (EMT) was observed in CCA, represented by the disrupted epithelial cell-cell junctions and mesenchymal characteristics [107-109]. Wnt/ β-catenin signaling is one of the critical pathways promoting the EMT transition [110,111]. In CCA cells, suppression of Wnt/β-catenin signaling increased E-cadherin and downregulated vimentin [112,113], suggesting that the Wnt/β-catenin signaling is associated with EMT during CCA tumorigenesis. β-catenin interacts with E-cadherin to form the cadherin-catenin-actin complex, maintaining epithelial cell adhesion, cytoskeleton, and integrity. During CCA development, the decreased E-cadherin releases β-catenin, resulting in β-catenin accumulation and nuclear translocation [111]. Then, β-catenin activates the transcription of twist, snails, and ZEB1 to induce the EMT process in CCA cells [111].
HB
HB is a rare malignant tumor found in infants and children [114]. The preclinical and clinical studies showed the hyperactivation of Wnt/β-catenin signaling in HB. In HB cases, β-catenin was found to be increased in the cytoplasm and nucleus of the tumor cells [115,116]. While CTNNB1 mutations are limited in the exon 3 in embryonal HB, the CTNNB1 mutations in fetal HB encompass exon 3 and 4 [117]. Meanwhile, missense, deletion, or insertion mutations in the AXIN1 gene were detected in 8% of HB cases [118].
MANIPULATING Wnt SIGNALING
Porcupine (PORCN)
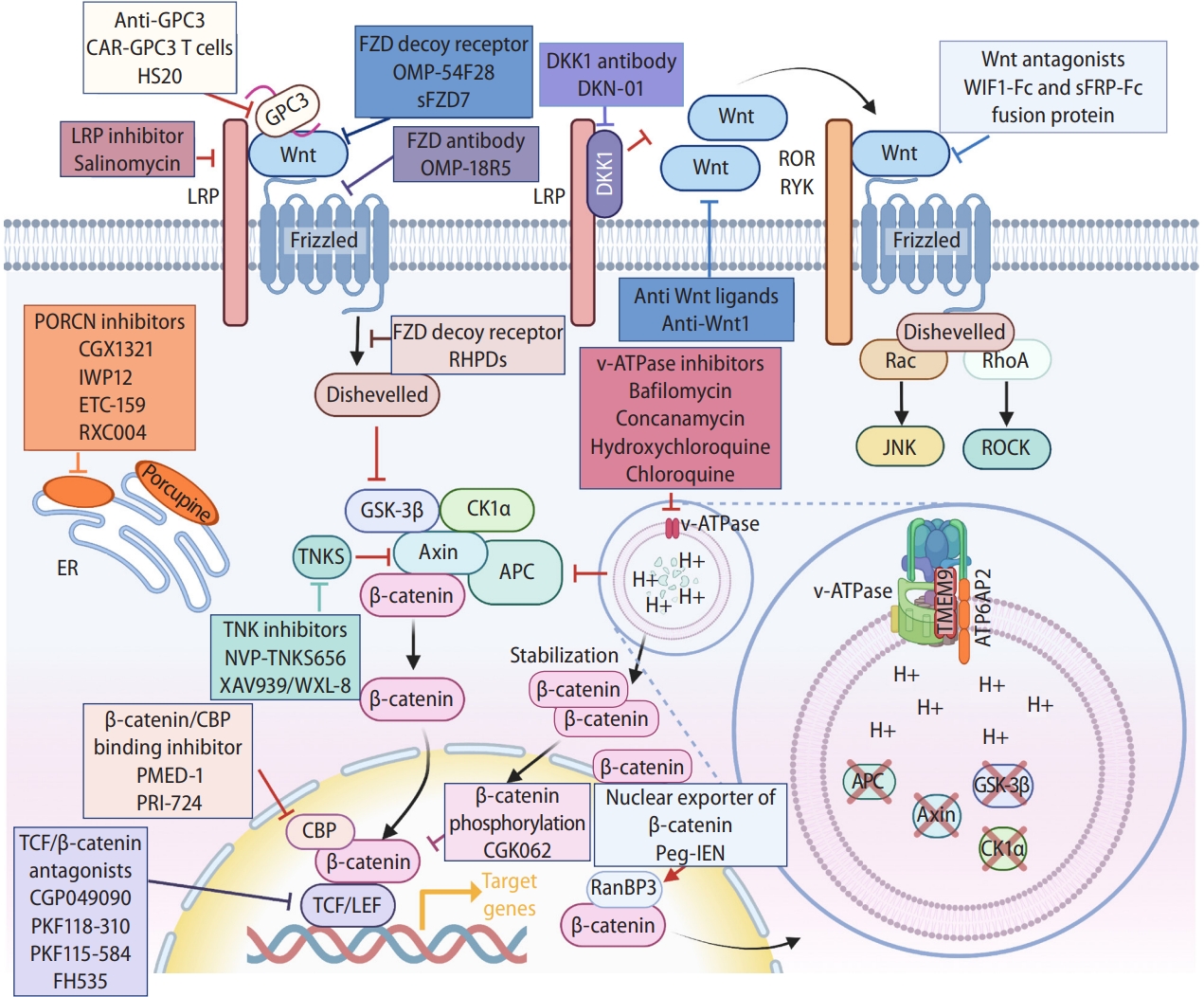
PORCN is a membranous protein mainly localized in the endoplasmic reticulum. PORCN mediates the palmitoylation of Wnt ligands, an essential process for Wnt ligands secretion and ligand-frizzled receptor binding [119-121]. Genetic and pharmacological blockade of PORCN reduces palmitoylation and inhibits the secretion of Wnt ligands, suppressing Wnt signaling [122]. The clinical trials showed promising results of PORCN inhibitors in HCC treatment. ETC159, CGX1321, and RXC004 have entered phase I clinical trials, and IWP12 is still in the preclinical studies (Fig. 4) [123]. In mouse models, Porcn KO induces embryonic lethality [124,125]. Porcn inhibition could cause adverse effects on bone homeostasis [126].
Wnt ligands
In physiological conditions, Wnt signaling is activated by binding of secreted Wnt ligands to LRP5/6 coreceptors and FZD receptors [127]. Thus, targeting Wnt ligands by chemicals or neutralizing antibodies efficiently inhibits Wnt signaling. Based on the high expression of WNT1 in human HCC cell lines and tissues. Anti-WNT1 neutralizing antibody showed its growth inhibitory effect on HCC cell lines but not on normal hepatocytes, with reduced β-catenin’s transcriptional activity (Fig. 4) [128].
Wnt antagonists
SFRPs, WIFs, and DKKs are the secreted Wnt signaling antagonists [129,130]. SFRP-1 and Wnt inhibitory factor 1 (WIF1) inhibit Wnt signaling by directly binding to Wnt ligands [131]. The fusion proteins WIF1-Fc and SFRP1-Fc were constructed by adding the Fc fragment of human immunoglobulin (Ig) G1 to WIF1 and SFRP1, respectively (Fig. 4) [132]. The fusion proteins exert potent anti-tumor activity by downregulating E2F transcription factor 1 (E2F1), cyclin D1, and c-Myc, increasing apoptosis of HCC cells and impairing tumor vascularization. DKK1 was initially considered a β-catenin-dependent tumor suppressor [130,133]. Several studies have shown that DKK1 promotes tumor cell proliferation, which may be due to DKK1-induced endocytosis of LRP and subsequent activation of the Wnt/PCP signaling pathway [134,135]. DKN-01 is a humanized monoclonal antibody targeting DKK1 in phase I/II clinical trial for HCC (Fig. 4). Phase I investigated the safety of DKN-01 as a single agent and in combination with sorafenib to treat HCC. Phase II explores the anti-tumor activity and safety of DKN-01 in patients with advanced HCC.
FZD receptors
The FZD receptors are promising therapeutic targets for HCC. The anti-FZD antibody can effectively reduce the HCC tumor growth by blocking the activation of FZD receptors on the Wnt signaling [136]. FZD decoy receptor OMP-54F28 (ipafricept) is a recombinant fusion protein that binds to a human IgG1 Fc fragment of FZD8 [137,138], which acts synergistically with chemotherapeutic agents (Fig. 4) [139]. A phase 1b dose-escalation clinical trial evaluated the safety, tolerability, and pharmacokinetics of OMP-54F28 when combined with sorafenib. Secreted FZD7 (sFZD7) is the extracellular domain of FZD7, expressed and purified from Escherichia coli. sFZD7 binding to WNT3 decreased the transcriptional activity of β-catenin/TCF4 and inhibited the growth of HepG2, Hep40, and Huh7 [140]. In combination with doxorubicin, sFZD7 inhibited the expression of c-Myc/MYC, Cyclin D1/CCND1, and Survivin/BIRC5, reduced the phosphorylation levels of AKT and ERK1/2, inhibited the growth of Huh7 xenograft tumors, and acted as a chemosensitizer [140]. OMP-54F28 is entering phase I clinical trials, while sFZD7 remains in preclinical studies (Fig. 4).
FZD antibody OMP-18R5 (vantictumab) is a monoclonal antibody directly binding to FZD receptors, which blocks the binding of Wnt ligands to FZD 1, 2, 5, 7, and 8 [141], which inhibits β-catenin-mediated transactivation (Fig. 4). In patient-derived xenograft models, OMP-18R5 combined with chemotherapeutic agents synergistically inhibited the development of several cancers.141,142 However, like PORCN inhibitors, OMP-18R5 has the same risk of impairing bone homeostasis [143]. In a dose-escalation clinical trial of OMP-18R5, one patient developed bone degeneration, controllable with zoledronic acid. The skeletal toxicity appeared to be manageable and reversible [144].
LRP co-receptors
Salinomycin (SAL), isolated from Streptomyces albus, is a monocarboxylic polyether ionophore antibiotic [145,146]. SAL blocks Wnt-induced LRP phosphorylation and leads to LRP protein degradation, destabilizing the Wnt/FZD/LRP complex and inhibiting the Wnt/β-catenin signaling (Fig. 4) [147]. SAL effectively inhibits β-catenin expression in HepG2/C3a cell line [148]. SAL also inhibits the migration and invasiveness of liver cancer stem cells through the Wnt/β-catenin signaling suppression [149].
Tankyrase (TNKS)
TNKS mediates PARsylation and subsequent degradation of AXIN via the ubiquitin-proteasome pathway, which in turn disrupts the β-catenin destruction complex [150]. Subsequently, the released β-catenin enters the nucleus to transactivate Wnt target genes [151,152]. TNKS is overexpressed in many cancers, including HCC, gastric cancer, and colorectal cancer [153-155]. The TNKS inhibitors XAV939, WXL-8, and NVP-TNKS656, attenuated Wnt/β-catenin signaling and inhibited the growth of HCC cells (Fig. 4) [155-157]. Moreover, TNKS inhibitors also suppressed HCC metastasis and invasion [157]. However, there are no relevant clinical trials for TNKS inhibitors in HCC.
Nuclear export of β-catenin
As shown in Table 1 and Supplementary Table 1, pegylated-Interferon-α2a (peg-IFN), the first-line therapy for the HCV-infected [158], attenuates the recurrence of HCC (Fig. 4) [159]. Mechanistically, peg-IFN upregulates the expression of Ran-binding protein 3 (RanBP3) [160], which enhances the nuclear export of β-catenin [160]. Thus, it is likely that peg-IFN-induced β-catenin nuclear export is a mechanism delaying HCC and improving survival in HCV patients.
β-catenin-mediated gene transactivation
The small molecule ICG-001 inhibits the interaction between β-catenin and CREB binding protein (CREBBP/CBP) for suppression of β-catenin-mediated gene transactivation (Fig. 4) [161]. A phase Ib/IIa clinical trial of the ICG-001 derivative, PRI-724, targeting HCC has been terminated [162]. Similar to ICG-001, PMED-1 disrupts β-catenin-CREBBP interaction and suppresses β-catenin target gene activation [163]. PMED-1 inhibits HCC cell proliferation but not normal human hepatocytes [163].
PKF118-310, PKF115-584, and CGP049090 are small-molecule inhibitors targeting the β-catenin-TCF complex (Fig. 4) [164]. These antagonists displayed the dose-dependent cytotoxicity in HepG2, Hep40, and Huh7 cell lines, with reduced cytotoxicity (10%) to normal hepatocytes. PKF118-310, PKF115- 584, and CGP049090 downregulated β-catenin target genes (MYC, CCND1, and Survivin/BIRC5) and inhibited the growth of HepG2 xenografts [164,165]. Similar to the mechanism of PKF118-310, PKF115-584, and CGP049090, FH535 inhibits β-catenin-mediated gene transactivation by interrupting the recruitment of nuclear receptor coactivator 2 (NCOA2)/GRIP1 to the β-catenin transcriptional complex [166]. It was shown that FH535 inhibits HCC cell proliferation by reducing cancer cell stemness [165].
Cancer-specific targeting of Wnt signaling
Given the pivotal role of Wnt signaling in the homeostasis and regeneration of multiple organs [168-170], broad-spectrum Wnt signaling inhibitors cause detrimental effects on the normal cells and organs. Therefore, cancer-specific Wnt signaling regulators may be attractive for Wnt signaling blockade therapy. TMEM9, an amplifier of Wnt/β-catenin signaling, promotes lysosomal protein degradation via v-ATPase, resulting in APC downregulation [46]. TMEM9 is highly expressed in liver regeneration and HCC. Genetic ablation of TMEM9 inhibits HCC tumorigenesis with downregulation of Wnt/β-catenin signaling [46]. Similarly, v-ATPase inhibitors, bafilomycin and concanamycin [171,172], also inhibit Wnt/β-catenin signaling without toxicity to normal cells and animals (Fig. 4, Table 1) [45,46]. Thus, molecular targeting of the TMEM9-v-ATPase axis can be used as cancer-specific Wnt/β-catenin blockade.
Glypican-3 (GPC3) is a proteoglycan binding to the FZD receptor and stimulates Wnt ligands-FZD interaction, resulting in the Wnt signaling activation (Fig. 4) [173]. GPC-3 is specifically expressed in HCC but not in normal human liver tissue [174]. The ectopic expression of GPC3 promotes the proliferation of HCC cells [175]. HS20 (an anti-GPC3 monoclonal antibody) suppresses Wnt/β-catenin signaling via inhibiting the interaction of Wnt3a with the GPC3 [176]. In xenograft mouse models, HS20 inhibited HCC progression without apparent concomitant toxicity [176]. To date, including CAR-GPC3 T cells or anti-GPC3 antibodies, 33 clinical trials related to GPC3 for HCC treatment were registered (https://clinicaltrials.gov/) (Table 1, Supplementary Table 1).
Concluding remarks
Wnt signaling activation plays a pivotal role in liver regeneration, metabolic zonation, liver diseases, and liver cancer. Aberrantly hyperactivated Wnt signaling promotes liver tumorigenesis and progression, often in conjunction with liver diseases. Although direct targeting of Wnt signaling sounds attractive as cancer therapy, given the crucial roles of Wnt signaling in tissue homeostasis and regeneration, severe adverse effects from Wnt blockade are inevitable. Nonetheless, an in-depth understanding of the biology of Wnt signaling in liver cancer and exploring cancer-specific Wnt signaling regulators are expected to identify molecular targets specific to liver cancer, which may overcome the current limitations of Wnt signaling inhibitors, and further improve therapeutic strategies of liver cancer treatment.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Supplement
Supplement Print
Print