INTRODUCTION
The liver receives a blood supply both from the portal vein and the hepatic artery. Portal venous blood, which is derived from the mesenteric veins, constitutes approximately 75% of total blood flow to the liver.1 Because large amount of microbes colonize in the gut, blood from the intestine contains not only products of digestion but also microbial products. Therefore, the liver, the initial site of filtration of gut-derived products, is susceptible to the exposure to the microbial products from the gut, such as lipopolysaccharide (LPS).1
In normal condition, translocation of the microbial products from the gut to extraintestinal space, including systemic circulation, is effectively prevented by our defense mechanisms: the barrier function of the gut and cleansing and detoxifying function of the liver.2 However, disruption of these defense mechanisms can lead bacterial translocation to extraintestinal space and aberrant activation of immune system, which can trigger harmful or chronic inflammations in the liver.3
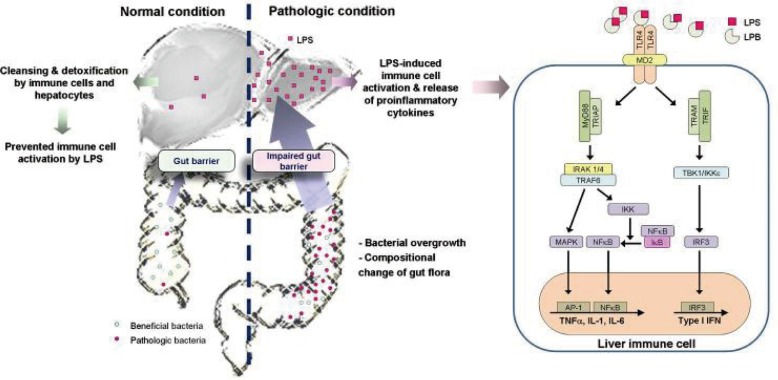
The importance of bacterial translocation in the pathogenesis of alcoholic liver disease has been shown in various previous studies: impairment of the function of intestinal tight junction4-7 and bacterial proliferation8,9 by alcohol and/or its metabolites, such as acetaldehyde, enhance bacterial translocation into the liver, which induce activation of immune cells, including Kupffer cells, to release various pro-inflammatory cytokines and chemokines (Fig. 1).10,11 Furthermore, various studies suggested that bacterial translocation also plays an important role in the development and progression of other types of liver diseases.12-16 Endotoxemia is frequently found in patients with cirrhosis, and the degree of endotoxemia is correlated with the degree of liver failure.17,18 In this review, we highlight the current knowledge about bacterial translocation and its contribution to the pathogenesis of chronic liver diseases and portal hypertension.
DEFENSE MECHANISMS IN THE BODY
LPS, glycolipids derived from the outer membrane of gram-negative bacteria, is a potent activator of immune responses: very tiny amount of LPS can induce the manifestations of septic shock in human.19,20 Fortunately, the mammals have effective defensive mechanisms to prevent this harmful effect of LPS. In healthy animals, LPS is cleared from the circulation within a few minutes after intravenous injection.21,22 Peripheral blood endotoxin concentration is significantly lower than portal venous endotoxin concentration, even in patients with liver cirrhosis.23
These defensive mechanisms mainly depend on the barrier functions of the gut and the detoxifying capacity of the liver.24 The gut epithelium acts as a first-line barrier to the gut microbes and prevents exposure of the gut microbes to the host immune system. The epithelial cells maintain barrier integrity by microvilli, tight junctions, and production of antimicrobial peptides.25-27 These barrier systems of intestinal epithelial cells prevent translocation of most of microbial products of gut and only tiny amount of microbial products can reach the liver in healthy condition.28
The liver plays a central role for prevention of translocation of gut-derived microbial products to the systemic circulation by cleansing and detoxifying microbial products.29 Microbial products in the portal venous blood eventually reach to the sinusoids in the liver, which contain diverse immunologically active cells, including the Kupffer cells, liver dendritic cells, T cells, natural killer T cells, and natural killer (NK) cells.28 Not only these immune cells, but also liver nonparenchymal cells such as hepatic stellate cells and liver sinusoidal endothelial cells, express the LPS receptor and remove this molecule to protect the systemic circulation from the endotoxemia.30 A previous animal study demonstrated that about 40-50% of intravenously administered LPS was quickly removed by the liver, suggesting the role of the hepatic uptake and detoxification in the immune homeostasis.31,32 Beside the LPS uptake, several other mechanisms also play a role in preventing significant immune response to LPS, including LPS-binding molecules, enzymes which degrade the lipid A moiety of LPS to decrease its activity, and LPS neutralization by serum lipoproteins.33-36 Furthermore, the liver usually tolerates bacterial products to avoid harmful responses.28 The hepatic immune system, including Kupffer cells, NK cells, NKT cells, T cells, and B cells, strictly regulate the liver immune system including liver tolerance.3
DISRUPTION OF GUT BARRIER FUNCTION AND THE DEVELOPMENT OF BACTERIAL TRANSLOCATION
It is well known that bacterial translocation is closely associated with the development of complications of liver cirrhosis, such as spontaneous bacterial peritonitis, the hyperdynamic circulatory state, and hepatic encephalopathy.37-39 Bacterial translocation is defined as the migration of bacteria or bacterial products from the gut to the extraintestinal space.40 Increased intestinal permeability induced by disruption of the intestinal epithelial barrier function contributes to the development of bacterial translocation.3,28,41 In addition, intestinal bacterial overgrowth and changes in the composition of bacterial flora in the gut can promote bacterial translocation.39,40,42
Increased gut permeability
The gut epithelium plays an important role in the immune homeostasis in the gut by acting as the first barrier against the bacterial translocation of gut microbiota.43-44 Because gut barrier system by intestinal epithelial cells prevent translocation of large amounts of bacteria and bacterial products from the gut, only very small amount of them can reach the liver in a healthy state.28 However, this effective gut barrier function can be disrupted by various pathological conditions and this disruption leads to bacterial translocation.3,28,41 For example, alcohol can play a role in the pathogenesis of alcoholic liver disease by disrupting the gut barrier function: alcohol itself as well as its metabolite, acetaldehyde, inhibit tight junction protein expression;44,45 alcohol can impair microtubule cytoskeleton in intestinal epithelial cells by inducing nitric oxide;46 and transepithelial electrical resistance is reduced in alcohol-exposed colon epithelial cells, which leads to impaired barrier function.24 In addition, liver-derived inflammatory cytokines can further increase gut permeability by disruption of gut epithelial tight junctions.47
Bacterial overgrowth
Bile acid secretion is decreased in patients with liver cirrhosis and this could lead bacterial overgrowth and compositional change in the intestine in these patients.48,49 In addition, the fact that liver cirrhosis and portal hypertension could impair intestinal motility may also contribute to the development of intestinal bacterial overgrowth.50 Previous study suggested that gastrointestinal transit is delayed in patients with liver cirrhosis and this delay could be improved with antibiotic therapy.51 Several studies suggested that the duration of the migrating motor complex are prolonged and the frequency of clustered contractions are increased in patients with liver cirrhosis, and these small intestinal motility disturbances are related with the severity of liver failure in these patients.52,53
Changes in the composition of gut flora
The composition of the intestinal bacteria is influenced by the environment, diet, and host factors.25,54,55 It could be changed by certain diseases, including liver cirrhosis. In liver cirrhosis, the normal intestinal microbial community is disrupted due to the decrease in gastric acidity, intestinal motility, and biliary secretions.56,57 Actually, a previous analysis of fecal microbiome in patients with cirrhosis suggested that fecal microbial communities are significantly different when compared to those in healthy individuals: increased prevalence of pathogenic bacteria, such as Enterobacteriaceae and Streptococcaceae, and decreased beneficial Bifidobacteria and Lachnospiraceae were noted in patients with liver cirrhosis.58,59 A previous animal study also reported the increased aerobic/anaerobic bacterial ratio in mice with liver fibrosis.60 In addition, liver cirrhosis induced by CCl4 in rats was also associated with high levels of Enterobacteriaceae.61 They also suggested that treatment with antibiotics or probiotics lead decrease in Enterobacter as well as increase in Bifidobacterium and Lactobacillus, which in turn lead to decreased systemic endotoxin levels and improve in the liver function.61 Similarly, treatment with probiotics or antibiotics in patients with liver cirrhosis reduced the prevalence of bacterial infection and hepatic enephalopathy62-64 and partially reversed the hyperdynamic circulatory state in these patients.65 A very recent study suggested that modulation of the intestinal microbiota is a critical determinant of nonalcoholic fatty liver disease (NAFLD) as well as multiple other aspects of metabolic syndrome.66
LPS RECEPTORS AND DOWNSTREAM SIGNALING PATHWAYS IN THE LIVER
In normal condition, only very small amount of bacteria or bacterial products, such as LPS, can enter the liver by the action of gut barrier function, where they are sensed and cleared by immune cells, particularly by Kupffer cells. However, in the pathologic condition with disrupted gut barrier function, the entry of bacteria and bacterial products to the liver is increased and homeostasis of the gut-liver axis is impaired, which eventually induce activation of liver immune cells, particularly Kupffer cells, to produce pro-inflammatory mediators.67-69 Previous studies suggested that bacterial translocation and resultant Kupffer cell activation are the main mechanisms of the pathogenesis of alcoholic liver disease.70 This suggestion is supported by the finding in animal models that alcoholic liver disease could be attenuated by gut sterilization with antibiotics or Kupffer cell elimination.71-73 Translocated bacterial products activate the hepatic immune cells through pattern recognition receptors, such as toll-like receptors (TLRs) and NOD-like receptors (NLRs). Recent studies suggested that TLR4 in hepatic stellate cells (HSCs) also responds to LPS to activate Jun N-terminal kinases (JNK) kinase and NF╬║B.74 Oxidative stress induced by alcohol and its metabolites is also involved in the induction of liver fibrosis by sensitizing HSC to LPS.75,76
Toll-like receptors
TLRs, a family of pattern-recognition receptors, are transmembrane proteins originally identified in mammals on the basis of their homology with Toll, a Drosophila receptor that contributes to development in the embryo, and the production of antimicrobial peptides against microorganism invasion in the adult fly.77,78 TLRs recognize pathogen-derived molecules-i.e., structural components unique to bacteria, fungi, and virus-and activate inflammatory responses including cytokine and type I interferon (IFN) production in response to this recognition.79 Previous studies suggested that hepatic non-immune cells, including HSCs and endothelial cells, respond to bacterial products through TLRs.3,28 Until now, ten TLRs have been identified in humans,80 while TLR4 was the first identified isoform that responds primarily to LPS.79
TLR4 plays a pivotal role in the activation of innate immune responses to LPS.81,82 TLR4 cannot directly bind to LPS and therefore, co-receptors, CD14 or MD-2, are needed for LPS binding to TLR4 and TLR4 activation.83-85 Two pathways for downstream signaling of TLR4 activation are demonstrated: MyD88-dependent and MyD88-independent pathways.86 In the MyD88-dependent signaling pathway, association of the intracellular TLR domain of TLR4 with the adapter molecule MyD86 through TRAM, which results in downstream activation of the IL-1 receptor-associated kinase 1 (IRAK1)/4/tumor necrosis factor receptor (TNFR)-associated factor 6 (TRAF6) complex and further activation of the I╬║B kinase (IKK) kinase complex that phosphorylates I╬║B to allow nuclear translocation of NF-╬║B.87 NF-╬║B binding to the NF-╬║B responsive element in the promoter region of pro-inflammatory cytokine genes results in the production of TNF╬▒, and other pro-inflammatory cytokines and chemokines.88 MAP kinases including c-JNK and p38 activate transcription factor AP-1. Activation of these transcription factors induces transcription of proinflammatory cytokines, such as TNF-╬▒, IL-6, and IL-1╬▓.89 Except for TLR3, all TLRs activate the MyD88-dependent pathway. In the MyD88-independent signaling pathway, recruitment of the adaptor TIR domain-containing adaptor inducing IFN-╬▓ (TRIF), TRAF6, to TLR4 and TANK-binding kinase (TBK)/IKK╬Ą phosphorylation induce phosphorylation of the interleukin regulatory factor 3 (IRF3), which in turn leads to IRF3 nuclear translocation and induction of type-I IFNs.88,90 Both MyD88-dependent and MyD88-independent pathways are activated after LPS-TLR4 interaction, while only one of these pathways are activated in other TLRs.
The importance of the TLR4 signaling pathway in the pathogenesis of alcoholic liver disease is evidenced by the previous animal study showing that decreased steatosis and inflammation and significantly reduced levels of pro-inflammatory cytokines, including serum TNF-╬▒ and IL-6, in the TLR4-deficient mice after chronic alcohol feeding.91 In addition, a previous study suggested that chronic alcohol exposure not only results in immune cell activation, but also sensitizes cells to LPS-induced pro-inflammatory signals by reduction in the expression of IRAK-M, a negative regulator of TLR4 activation.67 A critical role of LPS and TLR4 is suggested also in the pathogenesis of NAFLD: lipid accumulation, inflammation and fibrosis were significantly attenuated in TLR4 knockout mice after methionine choline-deficient diet.14,92
Recent studies suggested that TLR4 signaling can be activated not only by pathogen-associated molecular patterns (PAMPs), but also by some endogenous ligands, damage-associated molecular patterns (DAMPs), from cellular compartments which are released from damaged cells or tissues.79 DAMPs-induced TLR4 activation also can induce inflammation, which is called sterile inflammation because this inflammation is caused without infections.93 Therefore, DAMPs as well as PAMPs play a role in the pathogenesis and progression of liver diseases through activation of TLR signaling.3
NOD-like receptors and inflammasomes
NLRs is the members of the pattern recognition receptor family and they forms cytoplasmic multi-protein complexes, inflammasomes, with pro-caspase-1, the effector molecule, with or without the adapter molecule, such as the apoptosis-associated speck like CARD-domain containing protein (ASC).94-96 Inflammasomes are activated by sensing PAMPs or DAMPs via NLRs,97,98 and leads to activation of inactive pro-caspase-1 into active caspase-1, which in turn, induce cleavage of pro-inflammatory cytokines, including pro-interleukin (IL)-1╬▓ and pro-IL-18, into active forms of IL-1╬▓ and IL-18, respectively.94,99 IL-1╬▓ is a pro-inflammatory cytokine and plays a central role in regulation of inflammation by binding to the IL-1 receptor. IL-18 induces activation and IFN╬│ production of NK cells.100
CLINICAL SIGNIFICANCE IN CHRONIC LIVER DISEASES AND PORTAL HYPERTENSION
Alcohol can disrupt the intestinal epithelial cell tight junctions to impair the gut barrier function, which induce bacterial translocation and elevated endotoxin levels in the portal blood flow.105-107 Impaired gut permeability was also reported in NAFLD.12,16 A recent study suggested that modulation of the intestinal microbiota through multiple inflammasome components is a critical determinant of NAFLD/nonalcoholic steatohepatitis progression as well as multiple other aspects of metabolic syndrome such as weight gain and glucose homeostasis.66 In liver cirrhosis, the changes in intestinal motility and subsequent alteration of microflora content, decreased mucosal integrity, and suppressed immunity in hepatic fibrosis contribute to failure of the intestinal mucosal barrier with subsequent increases in bacterial translocation and LPS levels in hepatic fibrosis and cirrhosis.40,108-112
Consistently, plasma level of LPS is increased in patients with chronic liver diseases by viral hepatitis,31,112 alcohol,114,115 and NAFLD.116 Liver injury can be prevented by elimination of gram-negative microflora with Lactobacillus or antibiotics or Kupffer cell depletion with gadolinium chloride.71,72,117 Therefore, LPS-induced Kupffer cell activation is currently considered as a main mechanism for pathogenesis of alcoholic liver disease.118 Similarly, genetically obese rodents showed increased sensitivity to endotoxin119 and LPS challenge enhance the liver injury and induce inflammatory cytokine in mice with NAFLD.120 Some studies suggested that gut flora contribute to the pathogenesis of steatohepatitis in mice with fatty liver121 and treatment with probiotics reduce hepatic injury in obese mice.122 Furthermore, a previous study suggested that LPS-induced inflammation is involved in the pathogenesis of liver cirrhosis in patients with chronic viral hepatitis.123 Intraperitoneal administration of LPS can increase portal pressure124 and increased portal pressure can increase intestinal permeability.23,125,126 Bacterial translocation and increased production of proinflammatory cytokines and nitric oxide further impair contractility of mesenteric vessels in patients with cirrhosis, which could further increase portal pressure.127,128
A previous study reported that liver fibrosis and inflammation were significantly reduced after bile duct ligation in the TLR4-mutant mice and they suggested that LPS-TLR4 pathway play a crucial role in the hepatic fibrogenesis.129 It is supported by a large cohort study demonstrating that the TLR4 single nucleotide polymorphism predicts the risk of liver cirrhosis in patients with chronic hepatitis C infection.130
CONCLUSION
Various evidences suggest the gut-liver axis-from disruption of gut barrier function, bacterial translocation, and increase in LPS in the liver and systemic circulation to TLR and/or inflammasomes activation and production of proinflammatory cytokines-as the main mechanism of chronic liver disease and portal hypertension. Therefore, understanding this axis and the signaling pathway may provide new targets for the prevention or treatment of liver cirrhosis and portal hypertension.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print